Quanto � utile/interessante questa discussione:
| Autore |
Discussione |
|
|
netbridge85
Nuovo Arrivato

Prov.: Milano
Citt�: Milano

79 Messaggi |
 Inserito il - 08 luglio 2010 : 18:07:39 Inserito il - 08 luglio 2010 : 18:07:39




|
Tra le strategie utilizzate per identificare gli oncogeni nel genoma umano c'� quella di infettare le cellule (es. i fibroblasti)con il retrovirus. A questo punto gli LTR del virus si innestano ad es. vicino a geni protoncogeni e in seguito a riarrangiamenti il protooncogene diventa oncogeno che sotto il controllo del promotore virale LTR fa proliferare le cellule a manetta formando il tumore.
a questo punto dai foci ottenuti io prendo la cellula tumorale e sequenzio tutto il genoma per trovare i siti di inserzione degli LTR a fianco dei quali si troveranno tutti i geni potenzialmente oncogeni.
quindi vado ad identificare quali sono questi geni. La mia domanda sorge a questo punto. Non ho capito se in una stessa cellula infettata dal virus, l'LTR pu� piazzarsi una sola volta e quindi 1LTR per cellula e quindi un solo sito di integrazione per cellula, o se vi sono pi� LTR che si piazzano nel genoma della stessa cellula. Per� a logica suppongo che sia valida la prima...non tanto per il fatto che il virus ha un solo LTR, ma pi� che altro perch� se il mio scopo � quello di cercare gli oncogeni, e se pi� LTR si integrassero nel genoma della stessa cellula, sarebbe altamente improbabile che LTR che generalmente si piazza a caso nel genoma, vada a collocarsi nella stessa cell, accanto a tutti i protoncogeni.
Quindi visto che il mio dubbio ha a che fare con la tecnica, suppongo che io ho la mia bella piastra con i foci tumorali, in cui ogni clone (e quindi ogni focus nella piastra) rappresenti un LTR collocato accanto ad un dato protoncogene. quindi se "scansiono" tutti i genomi dei foci, trovo tutti i siti di integrazione (uno per clone) e mappo i geni posti a dianco dei siti di integrazione, trover� tutti i geni protoncogeni coinvolti nella formazione di tumore.
Qualcuno pu� dirmi dove e se sbaglio?
un altra domanda....ma l'LTR pu� controllare protoncogeni a grande distanza come gli enhancer? e quindi in ogni cell pu� controllare pi� geni contemporaneamente?
|
Se ti tramuti in qualcosa di pi� di un semplice uomo, se consacri te stesso a un ideale e se nessuno riesce a fermarti allora diventerai una leggenda! |
|
|
|
|
Dionysos
Moderatore


Citt�: Heidelberg
1913 Messaggi |
Inserito il - 08 luglio 2010 : 18:16:28

|
Non ho mai eseguito direttamente questo genere di saggi di trasformazione,
ma dato che lavoro in un laboratorio in cui si fanno anche queste cose
posso dirti che � possibile controllare la frequenza di integrazione
ed ottenere 1 popolazione a singola copia integrata. Ci sar� ovviamente
una deviazione (che segue la distribuzione di Poisson) ma selezionando
cloni ti sar� possibile isolare i singoli integranti, che sono gli
unici campioni biologici in grado di fornire un collegamento diretto
tra un preciso locus (quello che ha subito mutagenesi inserzionale)
e il suo diretto potere oncogenico - o meglio - trasformante.
Il potero oncogeno � una propriet� che si verifica in vivo:
iniettando le cellule trasformate in vivo, di solito in hosts
immunodeficienti, queste devono generare dei tumori spontaneamente. |
Volere libera : questa � la vera dottrina della volont� e della libert�
(F.W. Nietzsche)
Less Jim Morrison, more Sean Morrison!
|
 |
|
|
netbridge85
Nuovo Arrivato
Prov.: Milano
Citt�: Milano
79 Messaggi |
Inserito il - 08 luglio 2010 : 18:26:49
|
| quindi traducendo quello che hai detto vuol dire un LTR per genoma di una cellula, corretto? |
Se ti tramuti in qualcosa di pi� di un semplice uomo, se consacri te stesso a un ideale e se nessuno riesce a fermarti allora diventerai una leggenda! |
|
|
|
Dionysos
Moderatore
Citt�: Heidelberg
1913 Messaggi |
Inserito il - 08 luglio 2010 : 18:29:16
|
A rigor di logica due, se hanno una singola copia provirale
di retrovirus non self-inattivante |
Volere libera : questa � la vera dottrina della volont� e della libert�
(F.W. Nietzsche)
Less Jim Morrison, more Sean Morrison!
|
|
|
|
netbridge85
Nuovo Arrivato
Prov.: Milano
Citt�: Milano
79 Messaggi |
Inserito il - 08 luglio 2010 : 18:34:55
|
| si ma il secondo LTR in realt� corrisponde alle sequenze con i siti di terminazione della trascrizione, il poly-adenilation site etc...mentre il primo (a monte dei geni gag-pol-env)ha i siti per l'inizio della trascrizione.... |
Se ti tramuti in qualcosa di pi� di un semplice uomo, se consacri te stesso a un ideale e se nessuno riesce a fermarti allora diventerai una leggenda! |
|
|
|
Dionysos
Moderatore
Citt�: Heidelberg
1913 Messaggi |
Inserito il - 08 luglio 2010 : 18:51:38
|
... forse dovresti rivederti il ciclo replicativo dei retrovirus:
le LTR (long terminal repeats) sono chiamate cos� proprio perch�
sono sequenze ripetute in maniera eguale, sia a monte che a valle
del provirus. Questo perch� durante la retrotrascrizione (post-entry)
il virus replica la propria sequenza promotrice-enhancer (ossia U3)
in modo tale da anteporla al proprio transcription start site (TSS o +1).
Quindi ogni retrovirus avr� due sequenze U3 potenzialmente in grado
di disturbare la trascrizione dei geni adiacenti per cis-activation. |
Volere libera : questa � la vera dottrina della volont� e della libert�
(F.W. Nietzsche)
Less Jim Morrison, more Sean Morrison!
|
|
|
|
netbridge85
Nuovo Arrivato
Prov.: Milano
Citt�: Milano
79 Messaggi |
Inserito il - 08 luglio 2010 : 18:58:20
|
| uhm...non sapevo ci�...la mia prof ci ha detto cos�, come ti ho scritto su. adesso mi vado a rivedere un p� la question...grazie cmq. |
Se ti tramuti in qualcosa di pi� di un semplice uomo, se consacri te stesso a un ideale e se nessuno riesce a fermarti allora diventerai una leggenda! |
|
|
|
netbridge85
Nuovo Arrivato
Prov.: Milano
Citt�: Milano
79 Messaggi |
Inserito il - 08 luglio 2010 : 19:15:43
|
scusa dionysos, ho trovato questo pezzo (vedi testo in basso) in inglese che conferma la mia affermazione, e cio� che il 5' LTR agisce da promotore e che il 3' LTR da sito di terminazione, anche se sono uguali come sequenze. anche perch� altrimenti come fa il virus (o meglio la RNA pol II) a sapere dove finiscono i suoi geni? poi per� aggiunge, e questo valida la tua affermazione, che quando il 5' LTR � distrutto, allora il 3'LTR funge da promotore...quindi direi che sono giuste le posizioni di entrambi..no? :)
testo (link:http://www.stanford.edu/group/nolan/tutorials/retcl_3_ltrs.html)
The long terminal repeat (LTR) is the control center for gene expression. As may be expected because of the integrated phase of their life cycle, retroviruses have somewhat typical eucaryotic promoters with transcriptional enhancers and some also have regulatory elements responsive to either viral or specialized cellular (e.g. hormonal) trans-activating factors (HIV, MMTV). Enhancer functions have also been mapped to the gag (ALSV) and gag-pol (SIV and HIV) regions of some viruses but their role in the virus life cycle has yet to be established. In any case, all of the requisite signals for gene expression are found in the LTRs: Enhancer, promoter, transcription initiation (capping), transcription terminator and polyadenylation signal. Expression directed by the viral LTR signals is carried out entirely by host cell enzymes (RNA pol II, poly A synthetase, guanyl transferase). The enhancer and other transcription regulatory signals are contained in the U3 region of the 5' LTR, and the TATA box is located roughly 25 bp from the beginning of the R sequence. The presence and position of specific sequence signals used for transcription and binding sites for known or suspected transcription sites are shown in the figure below. The poly A addition signal (AAUAAA) in HTLV-1 is an unusually long distance upstream from the polyadenylation site (250 nt, by definition the end of R). It is hypothesized that the signal is brought in proximity to the addition site by secondary structure in the RNA.
The integrated provirus has two LTRs, and the 5' LTR normally acts as an RNA pol II promoter. The transcript begins, by definition, at the beginning of R, is capped, and proceeds through U5 and the rest of the provirus, usually terminating by the addition of a poly A tract just after the R sequence in the 3' LTR. Transcription is not terminated in the 5' LTR, probably due to failure of formation of secondary structure in the RNA which uncouples RNA polymerase, analogous to bacterial attenuation. Occasionally the stop signal in the 3' LTR is "read-through" and terminated in host flanking sequences, these hybrid transcripts could then be efficiently packaged and transmitted in virions. This process may play an important role in transduction of oncogenes.
The 3' LTR is not normally functional as a promoter, although it has exactly the same sequence arrangement as the 5' LTR. Instead, the 3' LTR acts in transcription termination and polyadenylation. Transcriptional interference occurs when the two LTRs are oriented as in a provirus; the 5' LTR has dominant control as a promoter. When the integrity of the 5' LTR is disrupted, the 3' LTR can act as a promoter, and this is the basis for one form of retroviral oncogenesis. Thus transcription from the 5' LTR interferes with the function of the 3' LTR as a promoter-enhancer. Experiments by Cullen et al. (Nature 307:239, 1984) demonstrated this phenomenon very clearly. By progressively removing portions of a provirus and using an indicator gene downstream of the 3' LTR, the transcriptional activity from both LTRs could be assessed. Cellular restriction of LTR function can occur. Transcriptional enhancers are tissue and cell specific in their function. For retroviruses like MuLV, embryonal tissues and cells do not host viral gene expression whereas differentiated or adult tissues and cells express viral RNA to high levels. These differences, which are due to enhancer "activators" and "repressors" have been studied in cell lines that can be differentiated in cell cultures. Changes in the structures of the binding sites in the LTRs can be correlated with tissue-specific expression patterns of mutant MLV and their induction of tumor formation in those tissues.
|
Se ti tramuti in qualcosa di pi� di un semplice uomo, se consacri te stesso a un ideale e se nessuno riesce a fermarti allora diventerai una leggenda! |
|
|
|
netbridge85
Nuovo Arrivato
Prov.: Milano
Citt�: Milano
79 Messaggi |
Inserito il - 08 luglio 2010 : 19:29:45
|
ho trovato anche un es. in cui il virus ha inizialmente i 2 LTR e poi perde per delezione quello a monte e funziona quello al 3' che agisce da promotore
Immagine:
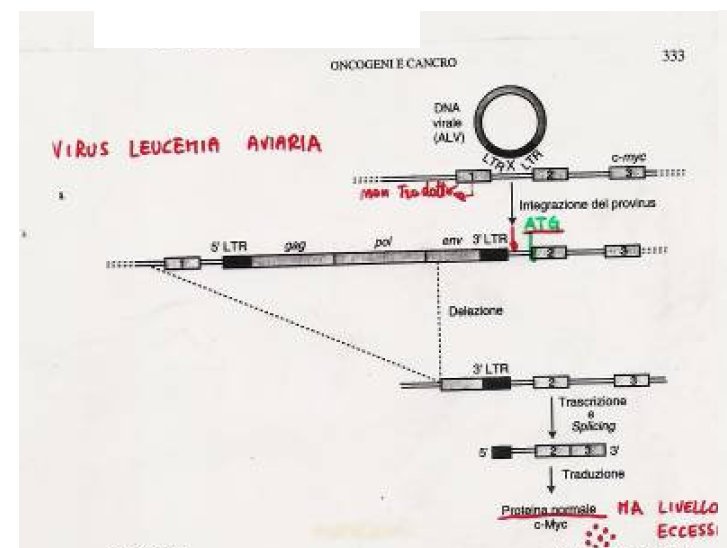
45,57 KB |
Se ti tramuti in qualcosa di pi� di un semplice uomo, se consacri te stesso a un ideale e se nessuno riesce a fermarti allora diventerai una leggenda! |
|
|
|
Dionysos
Moderatore
Citt�: Heidelberg
1913 Messaggi |
Inserito il - 09 luglio 2010 : 00:09:50
|
Infatti � contemporaneamente vero sia quello che dici che
che quello che dico io: non sono affatto incompatibili.
Domani ti spiego meglio, notte! |
Volere libera : questa � la vera dottrina della volont� e della libert�
(F.W. Nietzsche)
Less Jim Morrison, more Sean Morrison!
|
|
|
|
Dionysos
Moderatore
Citt�: Heidelberg
1913 Messaggi |
Inserito il - 09 luglio 2010 : 12:43:27
|
Dunque
la tua affermazione :
Citazione:
il 5' LTR agisce da promotore e il 3' LTR da sito di terminazione
� assolutamente corretta nella fisiologia del retrovirus:
la LTR (U3-R-U5) a monte � promotrice dela trascrizione dei
geni retrovirali, mentre la LTR (U3-R-U5) a valle agisce come
terminatrice. Entrambe le LTR posseggono sia regioni enhancer
e promoter (U3) che siti TSS o +1 (R) e pure un segnale di
poliadenilazione (tra R e U5) ma il funzionamento di questi
siti � context-dependent! quindi le cose stanno come dici tu.
Per� al di fuori della normale fisiologia di replicazione
del retrovirus, le due LTR sono esattamente identiche e
i sistemi cellulari che guidano la trascrizione dei geni
adiacenti il sito d'integrazione NON seguono lo stesso
schema trascrizionale del retrovirus: per questo entrambe
le regioni U3 verranno interpretate come enhancer/promoter
ed entrambe le regioni R-U5 come possibili siti di Poly-A.
A questo seguiranno fenomeni di cis-attivazione bilaterale
e di truncation trascrizionale del gene a monte (documentati).
|
Volere libera : questa � la vera dottrina della volont� e della libert�
(F.W. Nietzsche)
Less Jim Morrison, more Sean Morrison!
|
|
|
| |
Discussione |
|
|
|
Quanto � utile/interessante questa discussione:
| MolecularLab.it |
© 2003-24 MolecularLab.it |
|
|
|


