11 December 2010 - 22:08

Non capita tutti i giorni che gli scienziati si mettano a parlare di alieni. Ma di “alieni” invece se ne è parlato molto negli ultimi giorni, in seguito ad un annuncio da parte della NASA, la ben nota agenzia spaziale americana. In realtà, la parola alieni è stata utilizzata solo a scopo divulgativo, probabilmente per destare l’attenzione del pubblico. Pur avendo a che fare con una scoperta di un certo interesse, non si tratta in realtà di alcun essere venuto dallo spazio. La notizia diffusa dagli scienziati americani riguarda piuttosto un batterio, più terrestre che mai, in grado di sopravvivere sostituendo il fosforo con l’arsenico per il suo sostentamento.
La scoperta rientra in quella disciplina nota come astrobiologia (o esobiologia), una scienza che ha come oggetto di studio la vita al di fuori del pianeta Terra. Come tutti noi sappiamo, al momento non esistono forme di vita extraterrestri conosciute, di conseguenza l’astrobiologia non si occupa direttamente della vita su altri pianeti, ma piuttosto lavora formulando teorie su dove e come la vita potrebbe esistere in altre parti dell’universo. Gli astrobiologi elaborano i dati che le osservazioni spaziali forniscono, dati sulla presenza di pianeti intorno ad altre stelle, sulla loro distanza dal proprio sole, e sulla composizione dell’atmosfera. Queste informazioni servono a far capire agli scienziati se altri pianeti possano o meno avere le caratteristiche geofisiche e chimiche che sulla Terra hanno permesso la nascita della vita, ipotizzando di conseguenza se la vita possa essere presente anche su quei corpi celesti. In questo senso l’astrobiologia potrebbe essere considerata una scienza fondamentalmente speculativa.
Le recenti scoperte della NASA sono state presentate, prima dell’annuncio, come qualcosa che avrebbe dato nuova linfa alla scienza dell’astrobiologia. Non si sapeva ancora in cosa consistesse tale scoperta, ma come è facile immaginare l’annuncio ha presto fatto il giro del mondo ed è stato accolto come una possibile rivelazione della scoperta di vita extraterrestre.
Così non è stato. Non è stata trovata nessuna forma di vita aliena su Marte, su Titano o su qualche altro corpo extraterrestre. Quello che avevano scoperto gli scienziati della NASA era piuttosto un batterio terrestre, vivente nel Mono Lake, un lago salato californiano. Piuttosto vicino, si direbbe, rispetto ai pianeti extrasolari. Anche se vivente sulla Terra, questo batterio è stato comunque chiamato con l’appellativo “alieno”. Questo non tanto per il luogo dove vive, ma piuttosto per le condizioni in cui è in grado di vivere. Il microrganismo in questione è un ceppo degli Halomonadaceae chiamato GFAJ-1. Esso è in grado di crescere anche quando la concentrazione di fosforo è troppo bassa per permettere la vita, usando per il suo sostentamento l’arsenico, che è presente in concentrazioni relativamente elevate nell’ambiente in cui vive.
Come sappiamo il fosforo, sotto forma di fosfato, è un elemento necessario per la vita. Esso fa parte della struttura fondamentale degli acidi nucleici (DNA e RNA) ed è determinante per il funzionamento di proteine e lipidi. L’ATP, la molecola usata da tutte le cellule viventi come unità di scambio dell’energia, funziona proprio usando il legame altamente energetico fra gruppi fosfati.
L’ambiente dove vive GFAJ-1 è un lago salato che contiene alte concentrazioni di arsenico (200 μM). Gli scienziati hanno inserito un inoculo di sedimenti lacustri in un terreno contenente glucosio, vitamine e tracce di metalli, ma completamente privo di fosfato e additivi organici. L’unico fosfato presente nel mezzo derivava da impurità dei suoi componenti salini, ed era in concentrazione di solo 3,1 μM.
Addizionando arseniato in concentrazioni diverse i ricercatori sono riusciti ad isolare il ceppo in questione, che è risultato in grado di crescere nel terreno contenente il fosforo in basse concentrazioni e addizionato con l’arsenico. Per verificare che l’arseniato venisse effettivamente acquisito dal microrganismo è stata usata la spettrometria di massa. L’utilizzo di arseniato marcato con un isotopo radioattivo di As, inoltre, ha permesso di osservare la localizzazione intracellulare dell’arsenico utilizzato dalle cellule. L’arsenico è stato osservato in frazionamenti cellulari di proteine, metaboliti, lipidi e acidi nucleici. La presenza dell’arsenico nella struttura del DNA è stata in seguito verificata con ulteriori metodi spettroscopici, mentre la concentrazione di fosforo nel DNA dei batteri cresciuti nel terreno quasi privo dell’elemento sono risultate incompatibili con la vita.
Bisogna sottolineare che, tuttavia, il batterio non è un arsenofilo obbligato. GFAJ-1 cresce più velocemente quando gli si fornisce fosforo invece di arsenico, anche se il fosforo è di un ordine di grandezza meno concentrato dell’arsenico. Questo perché il fosforo dà luogo a molecole più stabili di quelle contenenti arsenio, e quindi è un elemento migliore per la costruzione delle biomolecole. Tuttavia gli esperimenti condotti dalla NASA hanno dimostrato che il microrganismo scoperto è in grado di sostituire il fosfato con l’arseniato nelle sue molecole biologiche nel caso il fosforo scarseggi. L’arseniato deve dunque poter adempiere a tutte quelle funzioni chimiche e dare luogo alle stesse interazioni del normale fosfato all’interno delle cellule.
Quali conclusioni scientifiche, e forse anche filosofiche, potremmo trarre da queste osservazioni? La scoperta della NASA non è importante in quanto conferma diretta di vita aliena, come molti speravano prima che i dettagli della scoperta venissero diffusi. Essa è probabilmente importante su un livello concettuale. Questo infatti è forse l’indizio più consistente del fatto che la vita, forse, può funzionare anche in assenza di tutte quelle condizioni che sulla Terra sono fondamentali per gli organismi, come la presenza di una sufficiente concentrazione di fosforo. Questa scoperta apre la strada ad un modo diverso di lavorare e ad ipotesi più flessibili riguardo la presenza di vita nell’universo.
Il fosforo è stato da sempre considerato un elemento di fondamentale importanza per il DNA e l’RNA, le molecole alla base della vita. Nonostante sappiamo da tempo che alcuni elementi accessori alla vita, come il ferro, possono essere sostituiti con altri, fino a poco tempo fa pochi biologi, probabilmente, avrebbero preso in considerazione che molecole come DNA e RNA potessero funzionare anche senza fosforo. Per quanto ne sapevamo, infatti, la composizione di tali molecole fondamentali è la stessa in tutti gli organismi conosciuti.
Si tratta questa di “un’altra vita”? Dobbiamo considerare GFAJ-1 un organismo a parte rispetto a tutti gli altri? La risposta credo che sia negativa. Nonostante la strategia usata dal batterio per sopravvivere sia per il momento unica, dobbiamo ammettere che tale caratteristica deve essere stata acquisita secondariamente durante l’evoluzione. Il batterio condivide antenati comuni a tutti gli altri organismi terrestri, antenati che si sono evoluti, probabilmente, con un DNA di struttura tipica. Dunque gli alieni ancora non sono scesi sulla terra e GFAJ-1 è più terrestre che mai. Ma questa scoperta dopo tutto ci porta ad una riflessione: forse la vita ha più assi nella manica di quello che pensavamo. Forse in futuro ci riserverà altre inaspettate sorprese che, perché no, potrebbero riguardare ambienti esterni al nostro pianeta.
Tags:
Arsenico,
Astrobiologia,
DNA,
GFAJ-1,
NASA
17 November 2010 - 18:13

I ricercatori hanno ottenuto cellule del sangue a partire dalla pelle
Negli scorsi articoli, parlando di possibili trattamenti per il ringiovanimento cellulare, abbiamo accennato alle iPSC, le cellule staminali pluripotenti indotte. Vediamo di approfondire l’argomento, parlando anche degli ultimissimi sviluppi che si stanno avendo in questo campo.
Ringiovanire le cellule
Le iPSC non sono né staminali embrionali né staminali adulte, ma sono ottenute attraverso la riprogrammazione di cellule mature, le quali sotto opportune manipolazioni regrediscono ritornando indifferenziate. Attualmente una dozzina di tipi cellulari (fra i quali la pelle) sono stati riprogrammati per trasformarli in staminali.
Per ottenere le iPSC gli scienziati hanno prima di tutto identificato quei geni che sono attivi nelle cellule pluripotenti e non in quelle differenziate. Di questi geni (circa una ventina) ne sono stati poi selezionati quattro che sono risultati quelli fondamentali per ottenere la riprogrammazione.
La messa a punto di questa tecnologia ha prima di tutto avuto delle implicazioni a livello concettuale: è stata di fatto smentita la convinzione che, una volta differenziata, una cellula non possa più tornare indietro allo stadio immaturo di staminale. Oltre a questo le staminali indotte hanno aperto anche nuove prospettive terapeutiche ed hanno risolto, almeno a livello teorico, molte delle problematiche legate all’applicazione terapeutica delle staminali “classiche”.
Quali sono i vantaggi che offrono le cellule staminali indotte rispetto alle tipiche cellule staminali embrionali o adulte? Anzitutto i problemi etici che spesso limitano gli scienziati nel lavorare con le cellule staminali embrionali non sussistono più con le iPSC, e questo non è poco. Le staminali indotte possono infatti essere fenotipicamente simili alle staminali pluripotenti ottenute da embrioni, ma di fatto esse non sono embrionali, cioè non si sono generate grazie ad un evento di fecondazione. Questa caratteristica le rende teoricamente immuni da discussioni di carattere etico o religioso, che nel nostro paese hanno un peso particolarmente elevato. Le iPSC sono cellule “qualunque”, che si erano differenziate, e che semplicemente sono state riprogrammate per tornare staminali. Insomma, cercare di riconoscere una dignità di “individuo” alle staminali indotte (come si fa con gli embrioni) sarebbe abbastanza forzato.
Oltre ai problemi etici con le iPSC sono stati risolte anche alcune difficoltà tecniche. Ottenere le iPSC è un processo più semplice a livello tecnico di quello che passa attraverso la clonazione per l’ottenimento di staminali embrionali.
Tuttavia la tecnologia delle staminali indotte è ancora acerba e necessita di perfezionamenti prima che possa essere sfruttata attraverso applicazioni pratiche. Le difficoltà, infatti, non sono ancora state del tutto superate. Sebbene le iPSC appaiano del tutto simili alle staminali pluripotenti normali ed esprimano i diversi marker di pluripotenza, non sempre riescono a comportarsi veramente da cellule pluripotenti. La ricerca è quindi attualmente concentrata a sviluppare metodi efficaci per riconoscere e selezionare le cellule staminali indotte in grado di funzionare bene dalle altre.
Cambio di identità
Mentre c’è che cerca di risolvere i problemi legati all’uso delle iPSC, non mancano altri sviluppi paralleli che riguardano il campo del controllo del differenziamento cellulare. Alcuni scienziati annunciano di essere già andati oltre questa tecnologia. Recentemente infatti la rivista Nature ha pubblicato un articolo nel quale alcuni ricercatori descrivono come siano riusciti a superare il concetto di staminale indotta, ottenendo cellule del sangue a partire da fibroblasti dermici senza passare per lo stadio di staminale. Grazie alle iPSC avevamo già capito che una cellula differenziata può tornare sui propri passi e rinunciare all’identità che aveva acquisito, diventando staminale e poi eventualmente differenziando in altri tipi di tessuti. Grazie a questo nuovo studio adesso sappiamo che lo stadio di staminale può essere scavalcato, ed un tessuto può essere indotto ad essere convertito in un altro direttamente.
I ricercatori della Mc Master University che hanno portato avanti questo studio sono riusciti quindi a cambiare l’identità delle cellule della pelle in quella delle cellule sanguigne. Questo è stato ottenuto attraverso l’induzione dell’espressione di fattori di trascrizione che di solito sono attivi durante l’ematopoiesi e grazie alla stimolazione con alcune citochine. Ciò ha portato prima alla generazione di cellule che esprimono il marker leucocitario CD45, e poi alle diverse cellule della linea leucocitaria ed eritrocitaria. L’aspetto importante è che tuttò ciò si è ottenuto per attivazione dei meccanismi adulti di ematopoiesi, e non di quelli delle cellule embrionali indifferenziate.
Dovrà passare ancora del tempo prima che si possa avere un’applicazione pratica e terapeutica di questi nuovi passi avanti della nostra conoscenza dei meccanismi di differenziamento cellulare. Sia le iPSC che le tecnologie che permettono il cambio diretto di identità cellulare sono ancora da perfezionare e ottimizzare. Questi risultati però arricchiscono sempre più la convinzione che lo sviluppo di biotecnologie simili sarà alla base della medicina di domani. I problemi etici derivanti dall’uso di cellule staminali embrionali potranno un giorno essere definitivamente scavalcati, e la cura di molte malattie potrà essere più semplice, più efficace e forse anche più economica.
Tags:
IPSC,
pelle,
sangue,
staminali
24 October 2010 - 15:38
Dal 1953, anno in cui Watson e Crick pubblicavano la struttura tridimensionale della doppia elica del DNA, sono stati fatti ampi progressi nel campo della genetica. Questi passi avanti hanno portato, negli ultimi 60 anni, a nuove incredibili conoscenze, dalla definizione degli RNA catalitici negli anni ’80 fino al completamento del progetto genoma umano alcuni anni fa. Nonostante questi grandi progressi, e nonostante oggi sequenziare un genoma sia diventato quasi una routine in alcuni laboratori, il sequenziamento dei genomi ci ha insegnato un’importante lezione: la conoscenza anche completa della sequenza nucleotidica rappresenta solo il primo passo nella scoperta delle funzioni del DNA, molecola che sembra riservare sempre nuove sorprese agli scienziati che si occupano di genetica.
L’era postgenomica
La differenza di sequenza fra il genoma umano e quello delle scimmie è estremamente bassa. Questo porta in evidenza un importante interrogativo: cosa rende diverse specie così simili per la sequenza del DNA? Se in passato si credeva che sequenziando il genoma di un organismo si potesse comprenderne tutti i processi biologici, oggi sappiamo invece che il pattern di espressione proteico di due cellule con contenuto genomico simile può essere molto diverso e la sua diversità non può essere compresa a pieno osservando la sola sequenza del DNA. Questo è il motivo per il quale, da qualche anno, è nata una nuova importante disciplina, chiamata proteomica. La proteomica ha lo scopo di studiare il cosiddetto proteoma, vale a dire l’insieme delle proteine espresse nelle cellule in un determinato momento o nel contesto di determinati processi. Lo scopo è anche quello di caratterizzare i meccanismi di regolazione che portano, da uno stesso genoma, alla definizione di quadri di espressione proteica anche molto diversi in tessuti differenti o in momenti diversi dello sviluppo. Si pensi al bruco e alla farfalla: stesso organismo e stesso DNA, ma fenotipo decisamente diverso.

Modello tridimensionale di RISC, complesso proteico implicato nel silenziamento a microRNA
I nuovi livelli della regolazione genica
Tali meccanismi di regolazione dell’espressione genica sono in parte ancora da scoprire. Se infatti sappiamo già molto sulla regolazione della trascrizione basata sull’interazione dei fattori di trascrizione con specifiche sequenze a monte o a valle dei geni, c’è ancora molto da scoprire quando si prendono in considerazione altri livelli sui quali può agire la regolazione dell’espressione genica. Ad esempio, la sintesi di una proteina può essere regolata a livello traduzionale agendo sul blocco o sulla degradazione del suo mRNA. Una strategia di questo tipo fa uso dei cosiddetti RNA antisenso. Essi sono molecole polinucleotidiche in grado di legarsi a mRNA complementari, impedendone così la traduzione da parte dei ribosomi.
I microRNA sono un altro tipo di RNA regolatori. Essi sono in grado di riconoscere mRNA a sequenza complementare e impedirne la traduzione guidandoli verso la degradazione. Sebbene i microRNA siano conosciuti da tempo e siano usati anche a scopo sperimentale per il silenziamento genico, recentemente una nuova scoperta ha reso il quadro ancora più complesso. Alcuni scienziati del Medical Center di Boston, infatti, hanno scoperto che il ruolo di microRNA e mRNA può essere ribaltato: RNA trascritti, ma non codificanti, possono legarsi ai microRNA impedendo che essi possano operare il silenziamento genico, aggiungendo così una altro punto alla lista dei possibili meccanismi di regolazione.
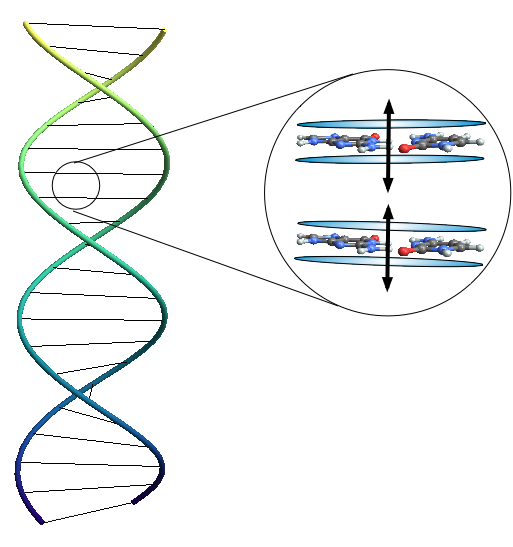
Applicando un modello di oscillatori armonici alla molecola del DNA, Elisabeth Rieper ne ha costruito un modello strutturale quantistico
La rivincita del DNA spazzatura
La scoperta di queste nuove molecole nucleotidiche implicate nella regolazione genica ha contribuito ad assegnare una funzione ad almeno una parte di quella porzione del genoma non codificante proteine, che negli anni passati era stata etichettata con il nome di junk DNA, vale a dire DNA spazzatura. Per molto si è discusso sulla possibile funzione di questa ampia porzione del genoma. Da molti era considerato nient’altro che DNA “egoista” che, simile ad un parassita, si era accumulato nei genomi pur non svolgendo alcuna funzione attiva, ma trasmettendosi di generazione in generazione insieme al DNA codificante. Altri ipotizzavano per il junk DNA una funzione di “tampone” per le mutazioni. Si era pensato cioè che il fatto che i geni fossero diluiti in mezzo ad una grande quantità di DNA inutile contribuisse ad abbassare la probabilità che una mutazione deleteria cadesse all’interno di un gene.
Oggi il DNA per il quale non si conosce una funzione sta diventando meno abbondante. Sempre più psuedogeni e sequenze di RNA non codificante vengono riconosciuti come responsabili della regolazione genica pur non sintetizzando proteine, ampliando così la porzione considerata funzionale del genoma.
Nuove frontiere: DNA quantistico
Una branca a dir poco avveniristica della biologia è la cosiddetta quantum biology, cioè “biologia quantistica”. Essa si prefigge di studiare l’implicazione dei fenomeni descritti dalla fisica quantistica all’interno dei sistemi biologici. Non accade spesso di sentir parlare di questo tipo di studi, ma i risultati ci sono e probabilmente ci aiuteranno in futuro a comprendere meglio i processi biochimici e la vita stessa.
Un fenomeno molto particolare del mondo dei quanti è il cosiddetto entanglement. Due particelle in stato di entanglement sono in grado, almeno apparentemente, di trasmettersi reciprocamente l’informazione sul proprio stato quantistico in maniera non-locale, cosa che da decenni assilla la mente dei fisici. Secondo uno studio condotto da Elisabeth Rieper della National University of Singapore il fenomeno dell’entanglement quantistico avrebbe un ruolo fondamentale nella strutturazione del DNA, affermando addirittura che secondo il modello strutturale classico la molecola del DNA non avrebbe energia sufficiente a rimanere unita.
Tutti questi nuovi dati e scoperte ci suggeriscono che il DNA, la molecola fondamentale della vita, racchiude ancora numerosi segreti che attendono di essere scoperti. Le osservazioni dei dati sperimentali, anche se ci forniscono nuove conoscenze, spesso introducono interrogativi nuovi, rendendo sempre più vario e complesso lo studio del fenomeno della vita.
Tags:
DNA,
entanglement,
junk DNA,
microRNA,
proteomica
9 October 2010 - 17:03
Alcuni esperti pensano che la medicina di oggi abbia raggiunto un livello sufficientemente avanzato da potersi dedicare, oltre alla cura delle malattie, anche alla battaglia contro il più endemico dei morbi: la vecchiaia.
Cerchiamo di fare il punto della situazione su quali siano i progetti e le speranze dei moderni scienziati.
Mangia meno e vivi di più?
Osservazioni compiute in vermi e moscerini hanno portato alcuni studiosi a trarre significative conclusioni: una dieta di restrizione calorica aumenta la longevità in questi animali, fino a raddoppiare la durata della loro vita. Questo principio potrebbe essere applicato anche all’uomo? Secondo Valter Longo, scienziato italiano all’estero che ha condotto studi di questo genere, probabilmente non è molto sano portare avanti un regime alimentare di restrizione calorica, ma si potrebbe fare qualcosa modificando il contenuto della dieta.
I risultati osservati negli animali potrebbero essere dovuti ad un fenomeno di adattamento evolutivo: il loro corpo in condizioni di basso apporto nutritivo non riesce a riprodursi in modo ottimale. È per questo che allora i meccanismi di invecchiamento cellulare verrebbero rallentati, in modo che l’individuo possa vivere di più per vedere tempi migliori, e sperare un giorno di nutrirsi sufficientemente per la riproduzione.
L’invecchiamento è regolato?
Queste supposizioni ci fanno intuire un punto importante: i fenomeni dell’invecchiamento sembrano essere regolati in qualche modo, dipendendo dagli agenti che operano sulle cellule e che influenzano le vie biochimiche responsabili. Se è così possiamo pensare, almeno da un punto di vista teorico, di poter agire su questi meccanismi di regolazione, in modo da manipolarli a nostro piacimento.
Secondo gli scienziati basterebbe addirittura modificare il regime alimentare, adottando ad esempio diete più povere di proteine, per riuscire ad allungare la vita.
Alcuni studi nel lievito hanno dimostrato, ad esempio, che esisterebbero delle vie di segnalazione in grado di agire sull’invecchiamento. Una di queste è il cosiddetto “bersaglio della rapamicina”. La rapamicina è una proteina immunosoppressore in grado di bloccare questa via. La sua somministrazione nei topi ha causato un aumento della longevità.
Ringiovanire le cellule
Quando si parla di sconfiggere la vecchiaia pensiamo subito all’invecchiamento dell’organismo in toto. Tuttavia, lasciate a parte tutte le aspirazioni di ottenere l’immortalità, potremmo pensare di agire solamente su determinate cellule e solo in situazioni patologiche, in modo da sfruttare il ringiovanimento cellulare solo a scopo terapeutico.
Grazie all’aggiunta di uno speciale miscuglio di geni che sono attivi solo negli embrioni, infatti, è stata dimostrata la possibilità di trasformare cellule adulte di topo in cellule staminali pluripotenti simili a quelle embrionali (le cosiddette cellule staminali pluripotenti indotte, o iPSC) aprendo la strada a possibili nuove terapie per combattere le malattie e gli acciacchi della vecchiaia.
Sono ancora molti i passi avanti da fare, ma se pensiamo che sono passati solo 13 anni dalla clonazione della pecora Dolly, i progressi scientifici in questo campo stanno procedendo ad un ritmo incalzante e le speranze per il futuro sono ben giustificate.
Le questioni etiche
Quando si parla di cellule staminali è inevitabile che, da qualche esponente politico o religioso vengano sollevate delle questioni etiche. Un esempio dell’ultim’ora è lo scalpore che ha suscitato in alcuni l’assegnazione del Nobel per la medicina all’inventore della fecondazione in vitro.
Cosa succederà quando si parlerà di allungare artificialmente la vita umana? Come reagirà l’opinione pubblica quando, inevitabilmente, gli scienziati cominceranno ad annunciare possibili terapie per vivere mille anni o più?
Difficile predire il futuro, ma si può dire per certo che argomenti così delicati non mancheranno di dare il via a nuovi dibattiti. E se ci guardiamo un po’ intorno, questo futuro sembra non essere neanche lontano, come ci dimostra quello che affermano gli scienziati che gestiscono Il Manhattan Beach Project, controverso progetto scientifico che promette di curare la vecchiaia entro il 2029.
Tags:
dieta,
invecchiamento,
IPSC,
staminali
10 April 2008 - 13:55
Uno dei delle domande esistenziali di una cellula è:
“Perchè una cellula dovrebbe suicidarsi?”
Non si tratta di masochismo cellulare, in certe circostanze la morte cellulare rappresenta un evento positivo per l’organismo.
Troviamo il fenomeno dell’apoptosi in un normale sviluppo embrionale, nella distruzione di cellule che rappresentano una minaccia per l’integritá dell’organismo; nel controllo della popolazione cellulare di un tessuto(omeostasi); nell’ eliminazione di cellule “Self” proprie “pericolose”
L’apoptosi è un sofisticato meccanismo in cui l’evoluzione cellulare ha fatto da setaccio per difendere l’organismo da cellule infettate da virus (ad opera di linfociti T citotossici); da cellule del sistema immune autoreattive; da cellule in cui si verifica un danno al DNA; da cellule tumorali.
L’Apoptosi è un processo attivo (richiede ATP) che viene indotto da segnali esogeni o endogeni e che serve ad eliminare, senza conseguenze cellule indesiderate. I processi fisiologici di regolazione dell’Apoptosi hanno segnali recettoriali, di danno direttamente al DNA e di segnali di membrana.
Se pensiamo allo sviluppo embrionale la morte cellulare è un processo fondamentale, per “scolpire” il nuovo organismo. Possiamo dire che la vita è un processo che in parte è reso possibile dalla morte. Infatti l’apoptosi durante lo sviluppo embrionale, per l’involuzione di strutture, può essere immaginata come una scultura di marmo; prima si parte dal blocco intero e poi piano piano si tolgono i pezzi con lo scalpello e il martello, così la scultura assume forma e sostanza. L’abbozzo delle mani è l’esempio più ecclatante per capire come funziona l’apoptosi nell’embriogenesi.
Altri segnali recettoriali di apoptosi sono nell’eliminazione dei neutrofilie macrofagi dopo l’infiammazione acuta, oppure nell’eliminazione dei linfociti T autoreattivi.
Per evitare l’accumolo di mutazioni che possono causare l’insorgenza di neoplasie, l’apoptosi gioca un ruolo fondamentale nell’eliminazione di cellule il cui DNA è stato danneggiato, cellule tumorali possono essere eliminate da parte dei linfociti T citotossici attraverso l’interazione di antigeni di membrana.
L’apoptosi é implementata mediante l’attivitá ordinata di
enzimi specifici, che agiscono prima della perdita dell’integritá della membrana cellulare.
Le caratteristiche morfologiche dell’apoptosi sono:
• Raggrinzimento cellulare;
• Condensazione e frammentazione della cromatina;
• Formazione di protuberanze (“blebs”) citoplasmatiche e di corpi apoptotici;
• Fagocitosi da parte di macrofagi o cellule non specializzate adiacenti
In vivo
– Le cellule “blebbing” sono ancora vitali (saggio del Trypan bleu);
– La transizione normale-raggrinzito-blebbing é rapida (10-30’), le cellule vengono poi fagocitate.
Istologicamente, l’apoptosi é quindi difficile da evidenziare a meno di un esteso coinvolgimento cellulare.
In vitro
Dopo il “blebbing”, c’é la vera e propria frammentazione cellulare in corpi apoptotici
• I corpi apoptotici vengono quindi fagocitati
• In situazioni -anche in vivo- in cui la apoptosi é massiva (es., nei tumori), cellule apoptotiche possono restare non fagocitate ed andare incontro a necrosi secondaria
In apoptosi c’è una progressiva degradazione proteica ad opera di proteasi specifiche (caspasi)
• Cross-linking proteico ad opera dellaTransglutaminasi II
• Frammentazione del DNA: formazione del “DNAladder”
• Attivazione di una “flippasi” e conseguente espressione di fosfatidilserina sulla parte esterna della membrana cellulare
Sperimentalmente si può evidenziare l’apoptosi di una determinata popolazione cellulare semplicemente osservando l’integrità del DNA
genomico per la presenza o assenza di attività endonucleasiche.

Perciò con un’ elettroforesi classica è possibile vedere i tagli delle endonucleasi mediate dalle caspasi che tagliano il DNA totale in frammenti di circa 200 paia di basi.
Line A DNA marker
Line B C Tipica strisciata di una cellula trattata con un apoptotico.
Linea D controllo Mock non trattato
Il metodo Tunel è una colorazione citochimica che sfrutta dei nucleotidi coniugati a digossigenina e aggiunti enzimaticamente al DNA ad opera della deoxynucleotidyl-transferase (TdT). Il DNA marcato viene rilevato grazie all’utilizzo di un anticorpo anti-digossigenina oppurtunamente coniugato con una molecola rivelatrice (fluorocromo, perossidasi ecc.ecc.)
L’apoptosi, è maggiormente resente in organismi cellulari, ma, in organismi monocellulari è presente in forme più o meno simili anche nei batteri, forse in difesa contro infezioni virali (a loro volta hanno escogitato numerosi stratagemmi per inattivare il programma apoptotico endocellulare)
Lo studio dell’apoptosi parte dal più famoso dei vermi trasparenti, il mitico Carnorabtis Elegans, durante lo sviluppo di C.Elegans, si formano 1090 cellule: 959resteranno a far parte del verme adulto e 131 vanno incontro ad apoptosi. Esperimenti di mutagenesi in cui il pattern di morte cellulare è alterato ha dato un grosso aiuto all’identificazione di regolazioni apoptotiche ritrovate anche nei mammiferi per analogia.

20 March 2008 - 22:17
La replicazione del DNA avviene tramite un modello discontinuo e semiconservativo cioè nel prodotto derivato, la molecola figlia, vi è sempre una molecola di sintesi e una proveniente dallo stampo.
Questo modello venne dimostrato da M. Meselson e W.F. Stahl:
I due ricercatori hanno coltivato Escherichia Coli in azoto 14 e 15 (+ pesante), attraverso combinazioni e aspettando la replicazione hanno successivamente estratto il DNA e il risultato era che la sequenza di bande rispettava il modello semiconservativo, avendo densità H-L
Proteine che intervengono nella replicazione
DNA polimerasi in cellule procariote, ne conosciamo tre:
•polimerizzano i nucleosidi trifosfati (crea nuovi filamenti)
•lavorano, solo in direzione5’ 3’, ma devono incontrare un DNA a doppia elica (anche 3’ 5’)
La più conosciuta è la III questa, ha 10 subunità:
• alfa(attività polimerasica), teta (attività esonucleasica), sigma(funzione non nota); tutte e tre rappresentano il CORE polimerasi
• beta, aumenta la processività, cioè il numero dei nucleotidi aggiunti prima che si stacchi del filamento, della DNA polimerasi
• tao, tiene legati due DNA polimerasi (punti core e subunità beta)
•complesso gamma, composto da 5 subunità attacca e stacca la subunità beta nelle fasi della replicazione
Quando entrambi i filamenti saranno replicati completamente, le DNA polimerasi 3 subiranno un distacco delle subunità beta, ma il core rimarrà attaccato per una successiva replicazione
•DNA elicasi e proteine stabilizzanti dell’elica (A e B), favoriscono l’apertura dell’elica da copiare
•DNA ligasi, tramite l’ATP unisce frammenti di DNA
•DNA topoisomerasi, favorisce la risoluzione dei problemi derivanti dall’avvolgimento dell’elica, (SUPERAVVOLGIMENTI).
•RNA primasi, aggiunge inneschi di RNA (primer, oligonucleotidi), si attacca alle proteine stabilizzanti l’elica.
Meccanismi che intervengono nella replicazione
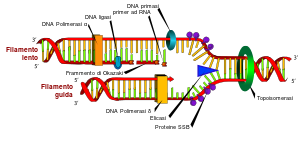

1) Svolgimento della doppia elica:
La proteine DNA A piglia contatto con la oriC, origine di replicazione, sulle sequenze ripetute 9 mer (basi) aprendo grazie all’ intervento di ATP la molecola di DNA nella 13 mer e costituendo così il complesso aperto; le regioni sono costituite di adenosina e timida (+ facili da scindere).
Sui due filamenti viene a posizionarsi di nuovo tramite una DNA C la elicasi (o DNA B), che svolge , idrolizzando l’ATP, il DNA procedendo in direzioni opposte dall’origine e formando il complesso di preavvio.
Le SSB (single strend binding, legano un filamento singolo), sono proteine che legano il DNA capaci di evitare il riappaiamento di due filamenti divisi attaccandosi su ognuno.
2)Replicazione nella forcella,: la forcella di replicazione ha forma di V con i suoi filamenti origine mentre quelli di nuova sintesi sono chiamati filamento guida (o veloce o leader), copiato grazie all’azione della DNA polimerasi dal filamento stampo (origine) 5’-3’ quindi avente direzione 3’-5’, e filamento in ritardo che siccome ha direzione opposta a quella in cui lavora la DNA polimerasi viene copiato attraverso un processo diverso ma che si ricongiunge a quello del filamento guida:
a.La RNA primasi aggiunge una serie di inneschi sul filamento in ritardo, lavorando in direzione 5’-3’
b.Si attaccano sull’estremità 3’ degli inneschi i frammenti di Okazaki
c.Gli inneschi sono rimossi dalla attività esonucleasica della DNA polimerasi 1, creando un GAP che poi viene riempito ,chiaramente partendo da 3’, dalla attività polimerizzante della stessa
d.Interviene la DNA ligasi che con l’uso di un a molecola di ATP realizza un legame fosfodiestere tra i frammenti di Okazaki
Se il GAP non fosse riempito, il filamento guida risulterebbe + corto del suo stampo di un lunghezza pari al suo innesco, e lo stesso problema si presenterebbe alla replicazione successiva. La molecola di DNA diventerebbe troppo corta per essere replicata e scomparirebbe dal genoma.
La lunghezza dei cromosomi è mantenuta costante da un blocco di sequenze ripetute in tandem che coprono le estremità, o telomeri di tutti gli enzimi eucariotici. Le telomerasi appunto aggiungono lr sequenze ripetu-te sul filamento stampo3’-5’.
Queste unità ripetute terminali sono tipicamente centinaia di copie di brevi sequenze ricche di basi azotate C e G, e si trovano nel filamento nucleotidico con l’estremità 3’ al telomero.
11 March 2008 - 01:18
Review

Per definizione è l’insieme di tutti i geni di un organismo e di tutte le sequenze intergeniche.
Il gene è quel tratto di DNA che codifica per la sintesi di un RNA o di una proteina
Nell’uomo ci sono 35-40000 geni che paragonati alla piccola negativa differenza di quantità dei nematodi (vermi) e dei pesci, e rapportando la complessità del nostro organismo con quello degli esempi ci si chiede quale sia il criterio che ci distingue.
La risposta è che la complessità del nostro organismo dipende dal prodotto (proteine) dei geni e non dal numero; infatti i geni si esprimono ognuno in maniera differente.
Si pensi anche che circa il 98% dei geni non si esprime, ma ha una funzione nella regolazione della stessa.
La quantità di DNA presente in un genoma (apolide) caratteristica di ogni specie è chiamata valore C , e la discrepanza tre grandezza gnomica e complessità genetica si chiama appunto paradosso C.
Il genoma è stato sequenziato nel 2001 da Collins e collaboratori, del NHRI (National Human Research In-stitute),scoprendo che il genoma è grande 3*10 alla nona bp (base pair = coppie di base, 3200 milioni di basi mb).
Il genoma sequenziato dell’uomo è di circa 2693 mb, e si divide in:
•25% GENICO
2,5% quello che si esprime
23% introni e pseudogeni
•75% INTRAGENICO
50% sequenze ripetute
24% duplicazione e altre sequenze
In definitiva il 97,5% dei geni del nostro corpo non si esprime, quindi rimane un 2,5% che sintetizza proteine. Comunque, quel 97,5% anche se non codifica il prodotto aiuta l’altra quota nell’espressione genica.
Ora, gli introni sono regioni non codificanti, gli esoni quelli codificanti, mentre gli pseudogeni rappresentano geni che non si esprimono (non sono attivi). Le sequenze ripetute sono principalmente divise in cinque classi:
a.Ripetizioni dovute a trasposoni (ripetizioni intersperse);
b.Ripetizioni di tipo (A)n, (CA)n, (CGG)n;
c.Duplicazioni (lunghezza 10-30000 kb)copiate da una regione all’altra del genoma;
d.Ripetizioni nei centromeri, estremità o intere braccia (cromosomi acrocentrici, cioè con braccia piccoline) chiamata RIPETIZIONE IN TANDEM;
e.Copie inattive di geni retrotrasposti, pseudogeni.
Analizzando la prima classe chiariamo che la trasposizione è un processo che permette lo spostamento del DNA mobile, costituente di una buona parte del genoma delle piante e degli animali superiori.
Il DNA mobile possiede due classi:
•Trasposoni
•Retrotrasposoni
Le differenze tra queste due classi risiedono nella differente modalità di inserimento della molecola di DNA:
il trasposone copia gli elementi direttamente in DNA,il retrotrasposone utilizza come intermedio una molecola di RNA cioè trascritte da una RNA polimerasi e converite di nuovo in DNA, per essere poi attaccate da nell’altra regione,da una trascrittasi inversa (questo processo, cioè il passaggio da una molecola di RNA ad una di DNA, nel 1975 non sembrava possibile).
RETROTRASPOSONI
•NON VIRALI
LINES
SINES (sequenze Alu)
•VIRALI
RIPETIZIONI LTR (Long Terminal Repeats)
I retrotrasposoni sono fonti di variabilità genetica, quindi anche di problemi in seguito a modificazioni non nella normalità.
Vengono distinti in:
Le ripetizioni LTR dei retroasposoni virali sono sequenze ripetute che permettono lo spostamento della molecola di DNA mobile, e rappresentano le porzioni terminali sia destra che sinistra della molecola. Al centro della stessa vi sono le proteine gag che hanno la funzione di codifica per la trascrittasi inversa.
Nei retroasposoni non virali, le “lines” sono sequenze ripetute di grandezza 6000-7000 bp (6-7 kb) e costi-tuiscono ben il 15% del genoma umano; le “sines”, 300-400 bp, sono principalmente rappresentate da se-quenze Alu cioè enzimi di restrizione che riconoscono la molecola di DNA e la tagliano.
LINE-Long Interdispersed Elements (6-7 kb)
SINE-Short Interdispersed Elements (300-400 bp)
3 March 2008 - 22:59
 Una delle domande che prima o poi tutti ci siamo posti conoscendo il mondo biochimico o biologico-molecolare; è :
Una delle domande che prima o poi tutti ci siamo posti conoscendo il mondo biochimico o biologico-molecolare; è :
come si fa a riconoscere il metodo di sintesi delle sostanze metaboliche o i metodi di degradazione e le cinetiche enzimatiche? Come facciamo a immaginare la forma dei cromosomi, a riconoscere il cromosoma 1 dal 2 ecc.
i metodi ovviamente non sono ipotizzati solo sulla carta, vero è che oggigiorno i metodi computazionali , informatici ci sono di grande aiuto, ma quando siamo completamente a digiuno di intermedi biomolecolari, quando non si sa da che parte incominciare; la “radioattività” ci dà una mano, come diceva tempo fa uno slogan a tutti noi molto noto.
Infatti il riconoscimento delle sostanze intermedie di una catalisi oppure di un processo metabolico o anabolico si conosce per brevi passi, effettuando dei piccoli approcci e unendo tassello dopo tassello (facendo delle radiografie intermedie) possiamo avere alla fine un quadro completo di tutto il nostro “percorso” metabolico.
Bè il caso vuole che le cose non sono così semplici come si direbbero, infatti per conoscere la via metabolica di una sostanza si utilizzano dei metodi poco invasivi per la cellula anche perché non devono essere perturbate le condizioni fisiologiche della cellula.
Uno dei metodi per conoscere una via metabolica è di introdurre un metabolita marcato con un isotopo quali ad esempio un C13, ad esempio può essere utilizzato un gruppo glucidico come il glucosio in cui un atomo di C è stato sostituito con uno marcato, successivamente si andranno ricercare le altre sostanze che presenteranno una attività radio, così abbiamo determinato i possibilili metaboliti del glucosio.
Tale metodica chiamata di diagnosi nucleare non si ferma solo all’individuazione dei processi metabolici, ma trova ampio impiego in medicina.
Un metodo ampiamente studiato e impiegato consiste nell’approccio con un chelante bifunzionale (Bifunctional Chelating Agent, BFCA). Non vorremmo parlare sempre delle stesse cose, ma il caso vuole che la chimica biologica, spesso, si avvale di questi metodi; il chelante bifunzionale presenta da un lato un set coordinativo in grado di stabilizzare un metallo radioattivo, dall’altro un gruppo funzionale per l’ancoraggio covalente della biomolecola, che può essere diretto oppure mediato da uno spaziatore (linker), a dare il derivato BFCA(-linker)-BM. La scelta accurata del BFCA è uno degli aspetti fondamentali nella progettazione di radiofarmaci target-specifici.
 In diagnostica ad esempio vengono utilizzati i cosiddetti radiofarmaci che sono delle sostanze radioattive in grado di legarsi e accumularsi sui tessuti bersaglio quali ad esempio organi interni come la tiroide e studiare, tramite visualizzazione di una lastra, la struttura dell’organo.
In diagnostica ad esempio vengono utilizzati i cosiddetti radiofarmaci che sono delle sostanze radioattive in grado di legarsi e accumularsi sui tessuti bersaglio quali ad esempio organi interni come la tiroide e studiare, tramite visualizzazione di una lastra, la struttura dell’organo.
Una dei nuovi approcci clinici in ambito di medicina nucleare si basa sull’uso di globuli bianchi marcati, i globuli bianchi come bene o male tutti sappiamo hanno funzione immunitaria, e quindi si accumulano in settori in cui è in atto un processo infiammatorio, ciò è utile alla diagnostica dello studio di processi infiammatori in atto, questa tecnica si chiama “scintigrafia”.
Viene ad esempio utilizzata quando si vogliono conoscere lesioni dell’apparato osseo, oppure dell’apparato nervoso.
Oppure è una tecnica utilizzata anche per diagnosticare se l’attività farmacologia antibiotica in atto è stata efficace.
Comunque a mio avviso una delle future applicazioni degli isotopi, è la diagnostica tumorale, in particolare delle metastasi tumorali, infatti utilizzando un trasportatore specifico possiamo evidenziare la location delle metastasi tramite scintigrafia, questo (come vedremo in seguito, nei prossimi post) è possibile, utilizzando dei trasportatori specifici che riconoscono delle cellule tumorali.
Tags:
analisi biomolecolare,
chelanti bifunzionali,
citologia,
immagini molecolari,
istologia,
scintigrafia
20 February 2008 - 18:19
Gli eucarioti sono cellule che hanno una struttura interna complicata ed un nucleo definito delimitato dalla membrana, e si differenziano da procarioti che presentano una struttura semplice e un nucleo ben definito. Il DNA è sempre associato a proteine che negli eucarioti prende il nome di cromatina, che è costitituita da:
Dna, istoni e proteine non istoniche
Il DNA è il contenitore del materiale genetico, avvolto ad opera degli istoni. Gli istoni sono proteine cariche positivamente, quasi sempre, a pH basico e costituite da lisina arginina e un gruppo fosfato che reagisce con le cellule vicine. Esistono 5 tipi di istoni:
- H1, chiamato di giunzione
- H2A
- H2B
- H3
- H4
E in ognuno di questi vi è una regione polare (con carica positiva, a seconda dell’intensità) e una centrale non polare, che possono modificare i loro amminoacidi o rimanere costanti.
Il 2, 3, 4, 5 presi due volte ciascuno formano un ottamero, che è avvolto da 1 ¾ digiro di DNA, pari a 46 basi azotate. Gli ottameri sono uniti tra loro dal DNA link (o connette) composto da 15 a 55 bp (coppie di basi). L’ottamero più il DNA link viene chiamato nucleosoma, unità fondamentale della cromatina, che de un aspetto di collana di perle. H1 e DNA link uniscono i nucleosomi (il filo che unisce le perle della collana).
 Gli istoni agiscono direttamente sul DNA principalmente come elementi strutturali mantenendolo in precise forme, modificabili con ubiquitina, gruppi fosfati e sostanze biologicamente attive.
Gli istoni agiscono direttamente sul DNA principalmente come elementi strutturali mantenendolo in precise forme, modificabili con ubiquitina, gruppi fosfati e sostanze biologicamente attive.
Le proteine non istoniche differiscono principalmente dagli istoni in quanto hanno una carica prevalentemente negativa, formano l’impalcatura della cromatina e posseggono la forma di un cromosoma metafasico (la metafase una fase della mitosi,abbiamo i cromosomi completamente condensati e attaccati all’equatore del fuso mitotico).
Le proteine non istoniche si dividono in 3 gruppi:
- Proteine che regolano la trascrizione genica.
- Enzimi e fattori attivi nella trascrizione, replicazione, ricombinazione, riparazione del DNA e modificazione del DNA di prot. cromosomiche.
- Proteine che contribuiscono al mantenimento della struttura della cromatina o della conversione della cromatina dallo stato decondensato allo stato altamente compatto.
Ricordiamo che il dna può essere distinto in due forme:
- CIRCOLARE esistente nei procarioti, mitocondri, cloroplasti, virus (un cromosoma ad anello associato a proteine, una origine di replicazione).
- LINEARE lo troviamo nei virus, sia Single Strend che Double Strend, eucarioti (più origini di replicazione).
La cromatinaè una struttura altamente dinamica infatti il nucleosoma subisce delle modificazioni strutturali durante la trascrizione e la replicazione: un rilassamento che avviene in tre processi:
- Acetilazione, in generale degli istoni (lisina) che riduce la condensazione della cromatina caricandola positivamente ed eliminando la sua interazione con un gruppo fosfato del DNA (attrazione elettrostatica)
- Scissione di H1
- Rottura dei nucleosomi tramite i complessi enzimatici della trascrizione e replicazione che legano il DNA con una maggiore affinità delle proteine istoniche
La cromatina è modificata anche nel suo spessore e nella sua forma:
- Nella forma: il DNA trascritto diviene una fibra spessa e quello non trascritto una collana di perle
- Nello spessore: la cromatina condensata diventa di 30 mm, mentre la non condensata prende la forma di una collana di perle di 10 mm.
La cromatina viene anche distinta in due tipi a seconda del suo avvolgimento:
- Eucromatina, forma meno condensata rispetto alla seconda, contiene geni sempre attivi nella trascrizione
- Eterocromatina, divisa in:
- Facoltativa, dove i geni possono esprimersi
- Costitutiva, nella quale l’espressione genica non avviene
Quando viene estratta dalle cellule in soluzione tampone isotoniche, la cromatina appare condensata (30mm) e i suoi nucleosomi (allo stato nativo) sono impacchettati in una forma a spirale chiamata solenoide. Il solenoide presenta da
6 a 8 nucleosomi per giro e l’H1 legato al suo interno.
Tags:
cromatina,
DNA,
istoni,
packaging
18 February 2008 - 20:58
 Visto i commenti che sono saltati fuori in merito ai composti chelanti, mi sembra giusto fare un po’ il punto della situazione con questi composti semplici funzionalmente ma dagli effetti fondamentali. Un tipico esempio di chelazione nei processi biochimici fisiologici è l’emoglobina, una proteina che contiene al suo interno dei gruppi –eme cioè delle strutture peptidiche (globina) contenenti, chelato, un atomo di ferro, e come tutti sappiamo la funzione del ferro legato alla globina e la formazione della emoglobina è il meccanismo fondamentale di trasporto dell’ossigeno alla cellula. Altro chelante endogeno, naturale, che la cellula da sola produce è la lattoferrina.La lattoferrina e’ una proteina chelante il ferro che e’ interessata in numerosi meccanismi biologici. È una proteina basica appartenente alla famiglia delle ferritine “non eme” cioè prive del gruppo eme, (polipeptide chelante il ferro), queste proteine, sono in grado di legare il ferro a PH neutro o alcalino e lo rilasciano a PH acido. Sono sintetizzate da particolari strutture esocrine come le cellule ghiandolari mammarie, e altri cellule secretive quali: lacrime, sudore, la bile, il liquido seminale e il succo pancreatico. Viene accumulata nei granulociti, e praticamente le ghiandole di tutti la classe dei mammiferi riescono a produrre questa proteina.
Visto i commenti che sono saltati fuori in merito ai composti chelanti, mi sembra giusto fare un po’ il punto della situazione con questi composti semplici funzionalmente ma dagli effetti fondamentali. Un tipico esempio di chelazione nei processi biochimici fisiologici è l’emoglobina, una proteina che contiene al suo interno dei gruppi –eme cioè delle strutture peptidiche (globina) contenenti, chelato, un atomo di ferro, e come tutti sappiamo la funzione del ferro legato alla globina e la formazione della emoglobina è il meccanismo fondamentale di trasporto dell’ossigeno alla cellula. Altro chelante endogeno, naturale, che la cellula da sola produce è la lattoferrina.La lattoferrina e’ una proteina chelante il ferro che e’ interessata in numerosi meccanismi biologici. È una proteina basica appartenente alla famiglia delle ferritine “non eme” cioè prive del gruppo eme, (polipeptide chelante il ferro), queste proteine, sono in grado di legare il ferro a PH neutro o alcalino e lo rilasciano a PH acido. Sono sintetizzate da particolari strutture esocrine come le cellule ghiandolari mammarie, e altri cellule secretive quali: lacrime, sudore, la bile, il liquido seminale e il succo pancreatico. Viene accumulata nei granulociti, e praticamente le ghiandole di tutti la classe dei mammiferi riescono a produrre questa proteina.
A questo punto corre in mente una domanda! ma perché la cellula produce lattoferrina? Cioè, qual è la funzione della lattoferrina? Perché il gene che codifica questa proteina si trova in particolare nelle cellule e ghiandole mammarie? Evidentemente visto che la molecola non contiene ferro, la sua funzione non sarà sicuramente quella di trasportarlo, o evidentemente di trasportare ossigeno. Anzi! La sua funzione è proprio quella di eliminare il ferro dal tessuto circostante nella quale si trova. Essa agisce legandosi e assorbendo il ferro, sostanza fondamentale per il nutrimento dei batteri; questi ultimi, privati del cofattore, muoiono. Questo meccanismo viene ormai utilizzato o sfruttato in farmacologia in quanto i batteri, privati dal ferro, sono costretti ad abbandonare le colonie, che spesso tendono a formare, diventando così più vulnerabili alle terapie farmacologiche. Infatti, i biofilm, hanno una massa batterica tale, da rendere inefficaci anche i più potenti antibiotici: questo ha sviluppato una tecnica farmacologica nell’integrazione o associazione degli antimetaboliti (antibiotici) con le lattoferrine.
Ma vediamo come è formata questa relativamente piccola, quanto importante proteina.  La struttura tridimensionale della lattoferrina umana, costituita da una singola catena polipeptidica di 692 aminoacidi, è organizzata in due lobi globulari, corrispondenti rispettivamente ai residui aminoacidici 1-333 (lobo N-terminale) e 345-692 (lobo C-terminale), uniti da una struttura ad alfa-elica comprendente i residui aminoacidici 334-344. Ciascuno dei due lobi della proteina è costituito da due domini. La proteina è strutturata in modo tale da disporre di un singolo sito di legame per il ferro per ogni lobo, situato in ciascuna delle facce interne dello spazio intra-dominiale. Le proprietà leganti della lattoferrina si sviluppano attraverso l’azione combinata di 4 residui aminoacidici identici per ciascun lobo (Asp58, Tyr93,Tyr193 e His254 nel lobo N-terminale, Asp396, Tyr434, Tyr527 e His596 nel lobo C-terminale) ed includono, oltre allo ione ferrico, un controione (normalmente un CO32-), sinergico per ogni Fe3+ e con la stessa attitudine ad accettare una gran varietà di cationi, in particolare Zn2+ e Cu2+ in luogo del ferro Questo set di ligandi è chimicamente e geometricamente ideale per un alta affinità di legame reversibile con il ferro e la presenza di un elemento non proteico (CO3 2-) sembra essere una condizione necessaria sia ai fini del legame, che del rilascio del ferro. Infatti, attraverso la neutralizzazione della carica positiva di un residuo di arginina (Arg121 nel lobo N) e della alfa-elica associata, l’anione CO32- facilita l’aggancio del ferro ai quattro ligandi raggruppati insieme sulla superficie laterale del dominio N2. Inoltre la protonazione dello stesso anione CO32- rappresenta probabilmente il primo step nel rilascio del ferro a bassi valori di pH.Una successiva protonazione della His in posizione 253, indebolisce ulteriormente il legame con lo ione ferrico, cui segue una dissociazione dei ligandi e una separazione dei domini, che realizzano il completo rilascio del ferro. Sito canonico di legame per il Fe3+ osservato nella lattoferrina. I due lobi della proteina nativa (non legata al ferro) possono assumere conformazione aperta o chiusa, e passare da una forma all’altra per effetto dei comparabili livelli energetici che caratterizzano i due stati. Generalmente le differenze nella struttura terziaria della proteina riguardano il lobo N-terminale, che nella apo-lattoferrina assume una conformazione aperta, mentre il lobo C-terminale resta chiuso. Probabilmente, il legame con il ferro si realizza quindi proprio nella forma aperta dell’apo-proteina, poiché essa consente un più agevole accesso agli ioni Fe3+ liberi o complessati. Coniugata al ferro si caratterizza invece per una conformazione chiusa dei due lobi.
La struttura tridimensionale della lattoferrina umana, costituita da una singola catena polipeptidica di 692 aminoacidi, è organizzata in due lobi globulari, corrispondenti rispettivamente ai residui aminoacidici 1-333 (lobo N-terminale) e 345-692 (lobo C-terminale), uniti da una struttura ad alfa-elica comprendente i residui aminoacidici 334-344. Ciascuno dei due lobi della proteina è costituito da due domini. La proteina è strutturata in modo tale da disporre di un singolo sito di legame per il ferro per ogni lobo, situato in ciascuna delle facce interne dello spazio intra-dominiale. Le proprietà leganti della lattoferrina si sviluppano attraverso l’azione combinata di 4 residui aminoacidici identici per ciascun lobo (Asp58, Tyr93,Tyr193 e His254 nel lobo N-terminale, Asp396, Tyr434, Tyr527 e His596 nel lobo C-terminale) ed includono, oltre allo ione ferrico, un controione (normalmente un CO32-), sinergico per ogni Fe3+ e con la stessa attitudine ad accettare una gran varietà di cationi, in particolare Zn2+ e Cu2+ in luogo del ferro Questo set di ligandi è chimicamente e geometricamente ideale per un alta affinità di legame reversibile con il ferro e la presenza di un elemento non proteico (CO3 2-) sembra essere una condizione necessaria sia ai fini del legame, che del rilascio del ferro. Infatti, attraverso la neutralizzazione della carica positiva di un residuo di arginina (Arg121 nel lobo N) e della alfa-elica associata, l’anione CO32- facilita l’aggancio del ferro ai quattro ligandi raggruppati insieme sulla superficie laterale del dominio N2. Inoltre la protonazione dello stesso anione CO32- rappresenta probabilmente il primo step nel rilascio del ferro a bassi valori di pH.Una successiva protonazione della His in posizione 253, indebolisce ulteriormente il legame con lo ione ferrico, cui segue una dissociazione dei ligandi e una separazione dei domini, che realizzano il completo rilascio del ferro. Sito canonico di legame per il Fe3+ osservato nella lattoferrina. I due lobi della proteina nativa (non legata al ferro) possono assumere conformazione aperta o chiusa, e passare da una forma all’altra per effetto dei comparabili livelli energetici che caratterizzano i due stati. Generalmente le differenze nella struttura terziaria della proteina riguardano il lobo N-terminale, che nella apo-lattoferrina assume una conformazione aperta, mentre il lobo C-terminale resta chiuso. Probabilmente, il legame con il ferro si realizza quindi proprio nella forma aperta dell’apo-proteina, poiché essa consente un più agevole accesso agli ioni Fe3+ liberi o complessati. Coniugata al ferro si caratterizza invece per una conformazione chiusa dei due lobi.
Attualmente si conoscono le sequenze aminoacidiche, determinate sia a livello proteico sia deducendole dalla sequenza nucleotidica, della lattoferrina umana, costituita da 692 aminoacidi. La lattoferrina umana possiede due potenziali siti di glicosilazione, uno in ciascun lobo. I residui glicosidici che contraddistinguono la proteina sono rappresentati essenzialmente da esossammine (fino all’1%) e zuccheri esosi (fino al 3%), in particolare mannosio comunemente legato ad un residuo terminale di fruttosio. L’elevato grado di glicosilazione proteica sembrerebbe attribuire alla lattoferrina una elevata resistenza all’attacco delle proteasi e all’abbassamento del pH, e sebbene possa significativamente influenzare l’interazione con molecole ligandi, la glicosilazione sembrerebbe non interferire con molte proprietà funzionali della lattoferrina. La concentrazione della lattoferrina varia ampiamente nelle diverse specie. In tutte le specie, comunque, tale concentrazione è più elevata nel colostro e può aumentare notevolmente durante una infezione intramammaria.
Tags:
antibiotici,
chelanti,
ferritina,
proteina

